

Главная >> Грибные статьи >> Что такое микориза
Что такое микориза?
Микориза, под этим названием известны соединения мицелия гриба с корнями высшего растения, причем последние при этом меняют более или менее характер своего роста, ветвления и анатомического строения. Микоризы очень широко распространены среди самых различных групп высших растений как семенных, так и архегониальных и, возможно, встречаются у большинства видов. По-видимому, совершенно лишены микориз представители Cruciferae, Polygonaceae, Cyperaceae, Equisetaceae. Среди других для некоторых образование микориз является обязательным (Orchidaceae, Ericaceae), а для большинства лишь факультативным, но все-таки и среди последних у некоторых, как у большинства наших лесных деревьев и многих преимущественно многолетних трав, микориза встречается особенно часто — в большинстве случаев. Разделяя растения по экологическим условиям, можно отметить, что микоризы всегда отсутствуют у водных растений и, наоборот, особенно широко распространены среди тех, которые растут на почвах, богатых гумусом. Далее, можно указать, что микоризы сравнительно редки у однолетних травянистых растений и часты у многолетних.
По строению различают эндотрофные и эктотрофные микоризы. У первых мицелий гриба распространен главным образом внутри тканей (в коровайларенхиме) и мало или даже в некоторых случаях совсем не выходит наружу. Корни при этом несут нормальные корневые волоски. У вторых мицелий оплетает корень снаружи, образуя около него как бы чехол грибной ткани с многочисленными свободными гифами, отходящими в почву. Собственных корневых волосков корень при этом не развивает. Переходом между этими двумя типами является так называемая экто-эндотрофная микориза, наблюдаемая, например, у наших берез и осин. Грибные гифы в ней не только густо оплетают корень снаружи, но и дают обильные ветви, проникающие внутрь в коровую паренхиму. Мицелий как типичной эндотрофной, так и экто-эидотрофной микоризы идет отчасти межклеточно, отчасти внутриклеточно. И тот, и другой в своих внутриклеточных частях образуют нередко густые клубки гиф или в других случаях — разветвления, напоминающие гаустории. Сами клетки корня при этом обыкновенно остаются живыми, и в некоторых из них, преимущественно более глубоко расположенных коровых клетках, наблюдается отмирание и растворение этих клубков мицелия. Это явление напоминает внутриклеточное переваривание и часто сравнивается с фагоцитозом у животных (переваривание микробов внутри лейкоцитов).
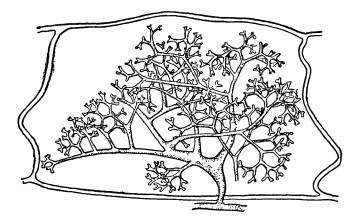
Типичная эндотрофная микориза свойственна, например, всем орхидным (за единственным, может быть, исключением Wullschlaegelia aphylla, совершенно лишенной микоризы). Присутствие гриба здесь является обязательным, особенно в молодых стадиях, а семена без инфекции их, как правило, совсем не прорастают (рис. 1).

Также обязательна эндотрофная микориза у представителей семейства Ericaceae. В подробностях она несколько отличается от того, что наблюдается у орхидных: она сосредоточена здесь главным образом в самых наружных эпидермальных клетках корня, где также образуются клубки, но переваривание их не наблюдается. Для ряда форм (Galluna, Andromeda, Vaccinium и др.) указывается, что гриб заражает сами семена уже в завязи. Таким образом, для их прорастания не требуется инфекции со стороны, что для орхидных, наоборот, является необходимым. Если искусственно освободить семена Galluna vulgaris от гриба, то хотя они и прорастают, но дальше нормального развития проростков не происходит (Rayner, 1915, 1929). Однако Knudson (1929) удалось получить нормальное развитие Galluna vulgaris в стерильных условиях.
Эндотрофная микориза встречается также у отдельных представителей самых разнообразных семейств цветковых растений (например, у Triticum, Zea, -Paris, Allium, Arum, Solanum, Nicotiana, Beta, Euphorbia и многих других). Ее присутствие не является здесь обязательным, и строение значительно отличается от вышеописанных. Во-первых, грибной мицелий здесь, как правило, неклеточный, тогда как у орхидных и вересковых он многоклеточный, во-вторых, он идет в более значительной части по межклетникам, в-третьих, внутриклеточные его части дают не клубки, а древовидные разветвления в виде гаусторий (так называемые арб у скул и — рис. 2) или на концах нередко пузыревидно вздуваются (так называемые спорангиоли).
Что касается, наконец, эктотрофной (и экто-эндотрофной) микоризы, то она представлена весьма типично у большинства, если не у всех, древесных пород, хотя и не является здесь безусловно обязательной.
Гриб в этом случае оплетает кончики корней, как бы чехлом, побуждая их вместе с тем к усиленному коралловидному ветвлению (рис. 3).
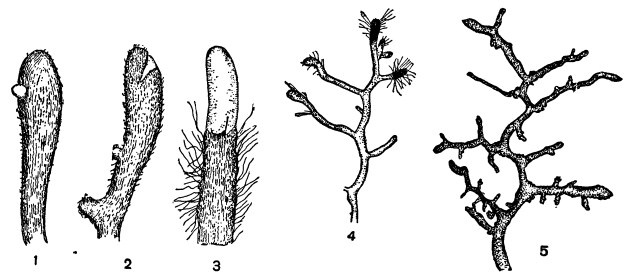
Наружные гифы указанного чехла свободно распространяются в почве, как бы заменяя отсутствующие здесь корневые волоски (рис. 3, 3, 4). Иногда при усиленном росте корня он разрывает на вершине грибной чехол и дальше растет свободно (рис. 3, 2, 3).
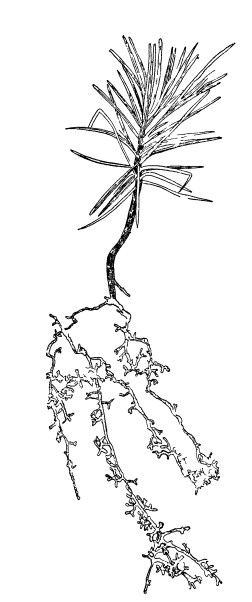
Анатомическое изучение микоризы наших древесных пород показывает ее экто-эндотрофный характер. Грибные гифы проникают между наружными клетками корня, особенно между клетками экзодермы, которые при этом значительно вытягиваются в радиальном направлении. Преимущественно однослойное грибное сплетение между этими и глубже лежащими клетками называется «сетью Гартига» (рис. 4).

Кроме межклеточного распространения, гифы проникают в самые клетки. При этом одни из них имеют вид толстых мешков, по-видимому, богатых питательными материалами, а другие— более тонкие и разветвленные. Они образуют клубни и нередко перевариваются в глубже лежащих клетках.
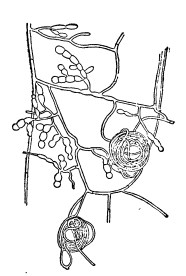
Грибы, находящиеся в микоризе, не образуют каких-либо органов размножения, по которым можно было бы непосредственно определить их систематическое положение. Для идентификации приходится поэтому прибегать здесь к косвенным и иногда сложным путям. Иногда самое прямое наблюдение в природе, показывающее постоянное нахождение данного вида гриба в ближайшем соседстве с данным высшим растением, имеющим микоризу, дает основание предполагать, что как раз между ними и существуют симбиотические микоризные соотношения. Более надежным является установление органической связи между микоризой и почвенным мицелием, образующим те или иные органы спороношения. Этот способ особенно применим и применяется по отношению к грибам с крупными плодовыми телами, как гименомицеты или трюфелевые. Однако и здесь он оказывается далеко не всегда осуществимым, так как проследить мицелий между частицами почвы — дело далеко не простое. Вернее, может вести к цели изолирование гриба из самой микоризы и изучение его в чистой культуре. Это было проделано планомерно в широком масштабе сначала Noel Bernard (1909) для микоризы орхидных, а затем многими другими авторами для других микориз. К сожалению, найденные грибы далеко не всегда дают в культуре органы спороношения, так что и здесь их точная идентификация трудна и даже иногда невозможна. Поэтому приходится применять еще другой способ: синтез микоризы из чистой культуры определенного гриба и стерильно выращенного высшего растения. В этом направлении работал с хорошими результатами особенно Melin (1921 до 1925). В результате весьма многочисленных наблюдений и опытов такого рода выясняется следующее: грибы орхидных в культуре обнаруживают большое сходство с грибом, весьма распространенным как в свободном состоянии, так и в виде паразита и известным под названием Rhizoctonia1 (рис. 5).
У японской бесхлорофилльной орхидеи Gastrodia elata микоризным симбионтом оказался опенок (Armillaria mellea). У Ericaceae и Pirolaceae в образовании микоризы принимают участие грибы, которые дают в культуре спороношения и оказываются принадлежащими к роду Phoma (из Fungi imperfecti, Sphaeropsidales), который вообще очень распространен как в сапрофитных, так и в паразитных условиях. Систематическое положение неклеточного мицелия в эндотрофных микоризах многих других растений остается неясным. Предполагают, что он принадлежит к фикомицетам, a Peyronel сближает его с Endogone В случае двойной инфекции вторым симбионтом считается также Rhizoctonia.
В образовании эктотрофной (и экто эндотрофной) микоризы древесных пород принимают участие весьма многие грибы, главным образом из гименомицетов, особенно из семейства Agariaceae, а также роды Boletus, Hydnum и некоторые другие.

Микоризы древесных пород оказываются гораздо менее специализированными. Например, у бука микориза может быть образована 12 различными гименомицетами и, кроме того, еще гастеромицетом, Scleroderma vulgare. Тоже у березы, лещины и многих других. С другой стороны, один и тот же гриб может давать микоризу с разными деревьями, например, Amanita muscaria — со многими лиственными и хвойными деревьями. Большую специализацию здесь показывают некоторые виды Suillus, особенно Suillus flavus, связанный исключительно с лиственницей. Также значительную специализацию имеет, по-видимому, масленок (Suillus granulatus, Suillus luteus) — с сосной, подберезовик обыкновенный (Leccinum scabrum) — главным образом с березой, и некоторые другие. Меньшим специалистом является, по-видимому, рыжик (Lactarius deliciosus), связанный с елью, сосной, пихтой и лиственницей.
Относительно физиологического характера взаимоотношений между обоими компонентами в микоризе так же, как и относительно лишайников, высказываются разные мнения. Несомненно, что и здесь существует борьба, и исход ее нередко определяется внешними условиями. Иногда микоризный гриб берет в ней перевес и как настоящий паразит убивает корень. С другой стороны, в некоторых клетках корня происходит переваривание гриба, а иногда оно доходит и до полного его уничтожения и освобождения корня от его сожителя. Однако и здесь, как в лишайниках, благодаря длинной истории совместной жизни оба компонента так взаимно приспособились друг к другу, что их сожительство в некоторых случаях сделалось для них обязательным, (облигатная микориза), и в нем получает известные преимущества для сохранения вида не только гриб, но и несущее его растение.
В свое время Шталем (1900) была высказана гипотеза о связи микоризообразования с транспирацией. Сущность ее заключается в следующем: перегнойные почвы, где особенно распространены микоризы, населены также огромным количеством микроорганизмов, в том числе и грибов. Поэтому возникает большая конкуренция между корнями высших растений и почвенными грибами в первую очередь из-за воды и минеральных веществ. В этих случаях эктотрофная микориза с мощно развитой системой отходящих от нее свободных гиф повышает поглощающую способность корня у тех растений, собственная корневая система которых развита недостаточно совершенно, и дает этим возможность выдерживать указанную конкуренцию. Однако и при этих условиях поглощающая способность таких растений остается, по-видимому, пониженной, так же, как и транспирация их. В связи с этим микоризные растения содержат сравнительно мало зольных веществ (в среднем около 5% по Шталю). Наоборот, другие растения, растущие на тех же гумусовых почвах, но обладающие более мощной корневой системой, достигающей тех горизонтов, где почвенные микроорганизмы уже не столь обильны, нормально развиваются без всяких микориз. Их поглощающая способность и транспирация стоят на более высоком уровне. Также значительно больше и содержание в них зольных веществ (10% и больше по Шталю).
Теория Шталя имела в виду главным образом травянистые растения и ставила упор на поглотительную способность корня и транспирацию. Исследования Мелина более глубоко освещают вопрос о значении эктотрофной микоризы у древесных растений. По его данным гриб здесь привлекается и стимулируется в росте какими-то выделениями корня (Melin предполагает фосфатиды). Они же способствуют и прорастанию спор микоризных грибов. Из ткани корня мицелий получает главным образом безазотистые органические вещества. С другой стороны, гриб своим густо разветвленным в почве мицелием, заменяя корню отсутствующие здесь у него корневые волоски, усваивает из почвы не только воду и минеральные соли, но и сложные, главным образом азотистые органические вещества (из лигнино-протеинового комплекса почвы). При переваривании гиф в клетках корня эти азотистые органические вещества идут на питание растения. Таким образом, оно через посредство гриба может использовать и те органические вещества почвы, которые прямо ему недоступны, а кроме того, конечно, воду и неорганические соли.
Что касается эндотрофной микоризы, то здесь физиологические отношения менее ясны. Установлено, что гриб здесь использует углеводы и другие, главным образом безазотистые, вещества корня, с другой стороны, наличие переваривания гиф указывает, что и корень должен получать что-то из гриба.
Остается недостаточно выясненным, что именно микоризный гриб воспринимает специально из окружающей среды. Предполагалось, что здесь в первую очередь имеет место усвоение свободного азота. Оказалось, что это имеет место не всегда, но в некоторых случаях это установлено экспериментально, например, для грибов из микоризы Ericaceae, относимых, как было сказано, к роду Phoma (Ternetz, 1906, Rayner, 1915). У них энергия усвоения азота оказалась в чистых культурах значительной, и экономический коэффициент (отношение усвоенного N к потребленному сахару) не уступающим в ряде случаев таким типичным азотособирателям, как Clostridium и Azotobacter (больше 1%). В связи с этим Ericaceae растут в почвах, бедных легко усваиваемыми азотистыми веществами.
Имеются также указания на усвоение свободного азота микоризообразователями орхидных (Wulf, 1927, 1933). Однако прирост азота, полученный автором в чистых культурах, был настолько незначительным, что его данные кажутся сомнительными. Некоторыми (Knudson, Freisleben) оспариваются, впрочем, и данные Rayner об усвоении свободного азота у Ericaceae. Однако здесь получалась в опытах достаточная прибыль азота, лежащая за пределами возможных неточностей методики.
Кроме возможного в некоторых случаях усвоения элементарного азота, эндотрофной микоризе нужно приписать еще и какие-то другие функции.
Весьма вероятно, что те, хотя и не особенно обильные, гифы, которые выходят наружу из эндотрофной микоризы, так же, как и в случае эктотрофной, воспринимают из почвы не только воду и соли, но и некоторые иначе трудно доступные органические вещества. Питание некоторых орхидных и Monotropa, лишенных хлорофилла и растущих в богатых гумусом почвах, настолько зависит от гриба, что некоторые авторы называют их даже не сапрофитами, как обычно, а паразитами, паразитирующими на своем грибе.
Наконец, в последнее время выдвигается положение о возможном значении микоризных грибов (как эндотрофных, так и эктотрофных) еще как поставщиков гормонов или биоса для тех растений, у которых собственная продукция этих веществ ослаблена или совсем отсутствует. В пользу такого представления можно привести особенно данные Burgeff (1934), который получил прорастание семян орхидных без участия живого гриба, в том случае, если он сеял их на убитые нагреванием культуры его на желатине. Такие же результаты получались от прибавления к культуре семян ацетоновых или спиртовых вытяжек из дрожжей. Burgeff прямо указывает, что в этих случаях дело сводится к действию витаминов (или бирса), так как редуцированные семена орхидей не содержат этих веществ и лишены способности сами их вырабатывать х. Возможно, что аналогичным образом следует объяснить результаты опытов Knudson (1924, 1929, 1933), который, в противоположность Rayner, получил прорастание семян Calluna и орхидных и дальнейшее развитие проростков без гриба при культуре на агаре с большим количеством органического материала. Также Freisleben (1934) указывает, что семена Vaccinium прорастают не только в присутствии своего микоризного гриба, но и других видов, как Penicilliuni, который никакой микоризы с ними не образует.
В заключение следует указать еще наперитрофную микоризу, понятие о которой недавно введено Jahn (1934, 1935). Он указывает, что у древесных растений в ближайшем соседстве с растущими концами их корней (в их так называемой ризосфере) находится довольно специальная флора грибов, различная для разных видов деревьев и для разных почв. Хотя они анатомически не связаны с корнем, но тем не менее имеют значение для его развития, создавая около него соответствующую среду. По мнению Яна, здесь имеет особое значение изменение реакции в области ризосферы, особенно в сторону подкисления, производимое грибами. Благодаря этому изменяется поглощающая способность корня и используются такие вещества почвы, которые иначе были бы недоступными для него. Вместе с тем перитрофная микориза представляет интерес как возможный первый этап эволюции микоризообразования, идущей далее к эктотрофной, экто-эндотрофной и, наконец, эндотрофной микоризе.
Разобранное выше явление микоризы, кроме своего микологического значения, представляет и более широкий общебиологический интерес. Из него видно, сколь многообразны и сложны могут быть соотношения в почве, которую нельзя рассматривать только как безжизненную массу минеральных частиц и растительных остатков на разных стадиях разложения. Благодаря обильному содержанию в ней микроорганизмов она приобретает гораздо более сложные свойства, как бы чего-то живого. Корни высшего растения, проникающие в такую густо заселенную почву, вступают в очень разнообразные отношения с ее населением. Конечно, в первую очередь здесь имеет место взаимная конкуренция, борьба за воду и питательные вещества. При этом корни сравнительно богатые органическими веществами, в том числе легко доступными углеводами, хемо-тропически привлекают к себе грибы и другие не зеленые микроорганизмы. На этой почве создается паразитизм многих грибов, которые совсем убивают и вытесняют корни. В других случаях дело не идет так далеко, и скопляющиеся около корня грибы, хотя и пользуются, надо думать, его выделениями, но не проникают в него. В результате естественного отбора наиболее жизнеспособными оказываются такие комбинации корня с окружающими его грибами, в которых корень не только не убивается, но и получает даже известные преимущества (например, в смысле указанного изменения реакции ризосферы). Получается перитрофная микориза. Отчасти из нее, а отчасти из паразитирующих на корнях грибов эволюционировали дальнейшие ступени микоризы. Будучи случайной и неуравновешенной на первых шагах эволюции, в дальнейшем под влиянием естественного отбора она совершенствовалась. Исчезали те комбинации, которые вели быструю гибель того или иного компонента, и оказывались более жизненными те, в которых, несмотря на взаимную борьбу, такой полной гибели не наступало. Последнее могло осуществляться в значительной степени благодаря использованию каждым компонентом некоторых противоположных функций другого: грибом — главным образом функции накопления углеводов в корне, а корнем — некоторых функций гриба, как его способность более полно использовать в условиях конкуренции с другими микроорганизмами не только водный и минеральный запас почвы, но и ее органические, в том числе азотистые, вещества. Как крайнюю ступень эволюции такого сожительства можно привести Monotropa и некоторые бесхлорофилльные орхидные (Neottia, Corallorhiza). Сюда же относится японская Gastrodia elata, интересная еще и тем, что микоризообразователем ее является опенок (Armillaria mellea). Все названные растения утеряли свой фотосинтез и перешли всецело на питание органическими веществами, которое получают из почвы через посредство сожительствующего гриба. В этом случае соотношения между компонентами перевернулись: высшее растение превратилось, так сказать, в паразита своего микоризного гриба.
Как пример другого рода крайнего приспособления можно привести злак Lolium temulentum. Органы этого растения всегда содержат грибные гифы. Содержатся они также и в семенах, обусловливая их ядовитое действие, сходное по симптомам с опьянением. Отсюда и все растение получило название: опьяняющий плевел. Для нас здесь существенно, что постоянное присутствие гриба и передача его из поколения в поколение не оказывает на растение угнетающего действия, хотя такое состояние опьяняющего плевела несомненно длится уже очень долго (по некоторым данным в семенах его, добытых из египетских гробниц и имеющих давность в несколько тысячелетий, обнаружено присутствие гриба). Несомненно, что в данном случае гриб питается за счет высшего растения, так как к фотосинтезу он, конечно, не способен. Однако эта отдача ему некоторого количества питательных материалов, очевидно, легко переносится растением, а какого-либо токсического действия на него гриб не оказывает. Некоторыми высказывается предположение о том, что гриб, может быть, ассимилирует свободный азот и этим оказывается полезным для плевела. Однако такое предположение ничем не доказывается и кажется мало вероятным. Всего вероятнее мы имеем здесь крайний пример взаимной приспособленности двух организмов, исходящий из первоначального паразитизма гриба на плевеле, или отношений, подобных микоризе, особенно таких, как у Calluna, где, по данным Rayner, гриб также переходит в надземную часть растения и заражает семена при развитии их в завязи.
Курсанов Л. И. Микология. 2-е изд. М., 1940. — 100-108 c.
По всем вопросам, связанными с работой сайта, обращайтесь на почтовый ящик администрации mushroom@tihaya-ohota.ru
Copyright ©2016 - 2024
